'These Institutions Are Betraying Their Own Values'—Here's How Offended Students Seized Power on Campus
At Middlebury, Claremont, Wellesley, and elsewhere, censorship is winning because faculty and administrators won't fight it.
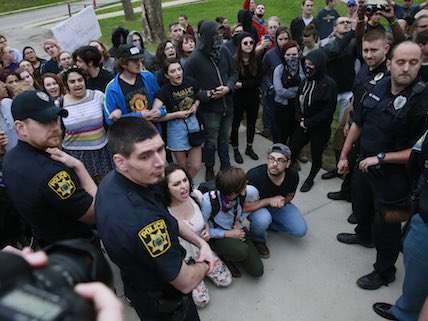
In the wake of the mob violence against Charles Murray at Middlebury College and Heather MacDonald at Claremont McKenna College, it's getting harder to deny that there is a censorship problem at American college campuses.
"These cases are proliferating," Nicholas Christakis, the former head of Yale University's Silliman College, told Reason in an exclusive interview.
Christakis, readers will recall, faced his own mob of unfriendly students a year and a half ago, after his wife—an early childhood educator—sent a campus-wide email pushing back against the administration's opposition to offensive Halloween costumes. The students implored Christakis to affirm that it was his job as an administrator to be their father figure on campus and provide a safe space where they would feel protected from emotional discomfort; when Christakis refused to do so—reasoning that learning is sometimes uncomfortable—they demanded his resignation.
The Yale incident happened in the fall 2015 semester. In retrospect, it almost looks like a high-water mark for the civil exchange of ideas. At least Christakis's students bothered to debate him: the mobs at Middlebury and Claremont McKenna so desperately wanted safety from offensive ideas that they were willing to resort to violence to get it. The 2016-2017 academic year has seen a transgender film director harassed at Reed College, windows smashed at the University of California-Berkeley, a controversial speaker maced at American University, and a liberal professors sent to the emergency room with a neck injury because she dared to have a conversation with Charles Murray.
Meanwhile, countless student groups, visiting speakers, and professors have had their rights infringed by administrators—sometimes at the behest of students.
"If you look at these incidents over the last couple of years, we're seeing escalations, in my view," said Christakis.
Indeed, the escalation theory is getting harder and harder to deny. While most college students remain friendly to freedom of expression and genuinely interested in hearing from speakers with different points of view, these students are increasingly cowed into silence by the small minority of censorious activists, New York University psychologist Jonathan Haidt recently told The Wall Street Journal.
"The great majority of college students want to learn," said Haidt. "They're perfectly reasonable, and they're uncomfortable with a lot of what's going on. But on each campus there are some true believers who have reoriented their lives around the fight against evil."
Nor is the "fight against evil" mentality reserved for use against characters like Milo Yiannopoulos, who are arguably beyond the pale. The Middlebury and Claremont McKenna incidents demonstrate that students are perfectly willing to shut down speakers who hold views that may be wrong, but nevertheless fall within the bounds of reasonable debate.
To prove this point—that Murray, an American Enterprise Institute and author of The Bell Curve and Coming Apart, is worth hearing out—Cornell University Professors Wendy Williams and Stephen Ceci recently surveyed dozens of professors for a New York Times piece. They sent the (presumably overwhelmingly liberal) professors a transcript of Murray's prepared remarks for his Middlebury lecture and asked them to rank the speech on a 1-9 scale in terms of how liberal or reactionary it was. Murray's talk received an average score of 5.05, indicating it was moderate in tone and substance.
That group wasn't told the remarks were Murray's. The second group was informed of the author's identity: this group gave Murray a score of 5.77.
"Our data-gathering exercise suggests that Mr. Murray's speech was neither offensive nor even particularly conservative," wrote Williams and Ceci. "It is not obvious, to put it mildly, that Middlebury students and faculty had a moral obligation to prevent Mr. Murray from airing these views in public."
What's not obvious to Williams and Ceci, Christakis, Haidt, or any number of other defenders of classically liberal values, seems very obvious to a whole host of people who make excuses for the students.
"There's nothing outrageous about stamping out bigoted speech," read the subhed of Slate writer Osita Nwanevu's piece responding to the Murray episode. The election of Donald Trump to the presidency has, in Nwanevu's view, dealt a blow to the idea that "those with the best command of facts and reason… will emerge victorious." The implication: the entire project of using reasoned debate to overcome harmful ideas in place of brute force might therefore be worth abandoning. How the worthy cause of halting Trump's anti-cosmopolitan agenda could possibly be served by a further retreat from liberal values is left unexplained by the author.
And yet Nwanevu's position is undeniably popular among the group of people for whom the benefits of free speech should seem most obvious: college professors. John Patrick Leary, an English professor at Wayne State University, celebrated that "Middlebury's students do, however, have every right to shout [Murray] down, and by all accounts they accomplished this end." Several professors at Wellesley College recently sent an email to students recommending that speakers like Laura Kipnis—a critic of Title IX witch hunts—no longer be invited to campus. The professors argued that people with disfavored opinions impose burdens on whichever groups are bothered by their views. The obvious flaw in this logic—what about the burdens imposed on the groups who want to hear the disfavored opinions?—was ignored, as is typical.
Wellesley's students apparently got the message. On Thursday, the college's student newspaper released a brazenly illiberal and poorly-written editorial that cheered on efforts to quash hate speech. No one on campus has a right to make statements that are sexist, racist, homophobic, transphobic, Islamophobic, or discriminatory in any way, according to the editorial. "If people are given the resources to learn and either continue to speak hate speech or refuse to adapt their beliefs, then hostility may be warranted," wrote the editors. What kind of hostility? You can guess. The undeniable subtext: Conform to ill-defined and constitutionally-suspect ideas about hate speech, or face violence.
In his interview with me, Christakis stressed that he shares students' commitments to fighting racism, sexism, and other societal evils.
"I am on the side of students," he said. "I believe in this generation. I believe in progressive values."
But censorship is not a progressive value, nor is violence. Christakis thinks professors are retreating from Enlightenment values and thus failing to instill proper liberalism in their students.
"I really believe the faculty, from an understandable position of empathy with the students, are nevertheless abrogating their duty to the students," he said. This has resulted in a failure on the part of colleges to foster appreciation for freedom of speech. "These institutions are betraying their own values," he said.
We are living with the results. In fact, Christakis is worried that one of the central tenets of the Enlightenment—that words are not the same thing as coercive violence—is being cast aside.
"During the Enlightenment we drew a big distinction between words and actions, and that the response to words is more words," he said. "We use our words so as not to fight. This was one of the great contributions of the Enlightenment. We used to burn people at the stake for saying the wrong thing."
Members of the faculty, particularly those who teach the humanities, social sciences, and the liberal arts, have an important role to play: they could reinforce the values of the institution, explain what a liberal arts education actually means, and push back against the ongoing conflation of words and action. And indeed, many continue to fulfill this role admirably. Not all campuses are plagued by censorship, and not all controversial speakers are run off campus. (Just last week, psychology professor Jordan Peterson, a critic of gender neutral pronouns, gave a talk at Harvard University that met with surprisingly little resistance.)
But more must be done to hold the line, because the power dynamics have shifted in favor of the most offended and speech-averse students at many of America's top colleges. As Murray himself noted, he has been speaking at university campuses for 20 years, and this was the first time the mob could not be silenced. At previous events, he wrote, "I could count on students who had wanted to listen to start yelling at the protesters after a certain point, 'Sit down and shut up, we want to hear what he has to say.' That kind of pushback had an effect. It reminded the protesters that they were a minority. I am assured by people at Middlebury that their protesters are a minority as well. But they are a minority that has intimidated the majority."
Take it from William Deresiewicz, author of Excellent Sheep: The Miseducation of the American Elite, who recently spent a semester at Scripps College and writes for The American Scholar:
I had one student, from a Chinese-American family, who informed me that the first thing she learned when she got to college was to keep quiet about her Christian faith and her non-feminist views about marriage. I had another student, a self-described "strong feminist," who told me that she tends to keep quiet about everything, because she never knows when she might say something that you're not supposed to. I had a third student, a junior, who wrote about a friend whom she had known since the beginning of college and who, she'd just discovered, went to church every Sunday. My student hadn't even been aware that her friend was religious. When she asked her why she had concealed this essential fact about herself, her friend replied, "Because I don't feel comfortable being out as a religious person here."
I also heard that the director of the writing center, a specialist in disability studies, was informing people that they couldn't use expressions like "that's a crazy idea" because they stigmatize the mentally ill. I heard a young woman tell me that she had been criticized by a fellow student for wearing moccasins—an act, she was informed, of cultural appropriation. I heard an adjunct instructor describe how a routine pedagogical conflict over something he had said in class had turned, when the student in question claimed to have felt "triggered," into, in his words, a bureaucratic "dumpster fire." He was careful now, he added, to avoid saying anything, or teaching anything, that might conceivably lead to trouble.
I listened to students—young women, again, who considered themselves strong feminists—talk about how they were afraid to speak freely among their peers, and how despite its notoriety as a platform for cyberbullying, they were grateful for YikYak, the social media app, because it allowed them to say anonymously what they couldn't say in their own name. Above all, I heard my students tell me that while they generally identified with the sentiments and norms that travel under the name of political correctness, they thought that it had simply gone too far—way too far. Everybody felt oppressed, as they put it, by the "PC police"—everybody, that is, except for those whom everybody else regarded as members of the PC police.
Permission to speak on campus is no longer absolute: it is managed and restricted by an unofficial ruling class consisting of a small number of students—the aforementioned PC police—who see no difference between speech and action, and reflexively lash out at any kind of expression that might offend someone. Professors are not taking strong enough action to fight this ruling class, and administrators are often complicit in its censorship.
And it's getting worse.




